 تويت
تويت
ملخص كتاب تصميم الحياة 7: التعقيد المحدد
المبحث الأول: التعقيد المحدد و علامات الذكاء
تظهر البنية تعقيدا محددا ان كانت لا يمكن انتاجها بالصدفة بسهولة و فى نفس الوقت تظهر نمطا محددا فمثلا سلسلة حروف مرتبة عشوائيا قد يعتبر أمرا معقدا و لكنه ليس محدد و بلا معنى أو وظيفة أو حتى نمط ظاهر بينما سلسلة حروف مرتبة لتعطى كلمة هو أمر محدد له معنى لكنه ليس معقد فقد يكون صدفويا (ومن هذا الأمثلة الداروينية العجيبة عن حبات الرمال و ندف الثلج المرتبة فى نمط بالصدفة ثم الاستدلال بها على امكان المصانع الخلوية و الالات البروتينية و مولدات الطاقة الجزيئية فى الخلايا و الكم الهائل من المعلومات الموجود فى الحمض النووى!!!) أما سلسلة مرتبة لتعطى نصا كامل المعنى فهى عملية معقدة و محددة لذلك يجب عزوها للذكاء.
https://billdembski.com/documents/2005.06.Specification.pdf
تستخدم الكثير من العلوم فكرة التعقيد المحدد كالطب الشرعى و علم الشفرات و الاثار و محاولات رصد اشارات من خارج الأرض. يستخدمأنصار التصميم الذكى نفس المبدأ مع النظم الطبيعية و الالات الجزيئية المعقدة للتدليل على أنها منتجات ذكاء و تصميم و يخضعون التصميم الذكى لمعايير رياضية صارمة ثلاث:
1-العنصر الاحتمالى: و يقيس احتمال حدوث التعقيد فمثلا عند توليد رقم عشوائى من 0 الى 9 5 مرات متتالية سيكون احتمال كل رقم أن يتولد 5 مرات متتالية 1 من 100000 و كلما كان هذا الرقم أكبر كان احتمال توليد النتيجة المطلوبة أقل و يصبح تفسيرها صدفويا أصعب
2-العنصر الوصفى للتعقيد و هو الذى يساعد على تحديد هل نحن أمام نموذج أم أمام عشوائية فمثلا أنت عندما ترى محرك ستوصفه ببساطة بأنه محرك برغم تكونه من أجزاء كثيرة متناسقة بينما اذا كانت كومة عشوائية من الأجزاء فلن تملك الا توصيفها بكلام كثير على غرار "قطعة معدن مستطيلة منثنية النهاية تدخل نهايتها فى دائرة معدنية ذات حواف بارزة...الخ". كلما كان الوصف أسهل فىالتعبير عنه و غير معقد دل هذا على وجود نموذج محدد و العكس يدل على العشوائية
3-المقياس الاحتمالى [حوض الاحتمالات]: و هو عدد الفرص المتاحة لانتاج الحدث فمثلا فى مثال الأرقام المذكور فى شرح العنصرالاحتمالى اذا قمنا بتجربة توليد الأرقام 100 ألف مرة فنحن قد ولدنا عدد من الفرص [حوض احتمالى] مناسب لانتاج النتيجة 1 من 100ألف بينما لو قمنا بالعملية عشر مرات فقط فان النتيجة تبدو بعيدة الاحتمال جدا فكلما تناقص هذا المقياس كانت الاحتمالات الصدفية اقل.
تعتمد الالية الداروينية الحديثة فى توليد الصفات الجديدة ليعمل عليها الانتخاب الطبيعى على الطفرات و أخطاء نسخ المادة الوراثية لذلك تلعب الاحتمالات فيها دورا مفصليا و هو ما يظهر أهمية المقاييس الرياضية فى تفنيد مزاعمها الى جانب العقل و المنطق و البداهة كما فى طرح مايكل بيهى عن التعقيد غير القابل للاختزال. يتبقى تشكيك الدراونة فى العنصر الوصفى فى أنه تحايل لغوى للايحاء بوجود تصميم أو كما يقول الداروينى مايكل شيرمر "نحن نتوهم وجود تصميم لأن الداروينية صممتنا لالتقاط الأنماط من البيئة حولنا" [و الحقيقةأن هذا اعتراف ضمنى بوجود أنماط فى البيئة و أن التقاطها و فهمها و عدم تجاهلها مهم للبقاء و الاستمرار – كما أن هذا الاعتراض لا يتفق مع اصرار الالحاد على انكار الفطرة الايمانية فقط من أجل الفطرة الداروينية] و الحقيقة أن الرد العلمى الواضح على هذا الاعتراض هو أن المصطلحات اللغوية المستخدمة فى توصيف الاليات الجزيئية و البروتينية هى مصطلحات هندسية نستخدمها لتوصيف الات صنعها البشر قبل أن يكتشف علماء الأحياء وجود نظائرها فى الخلايا و الكائنات فهل نحن ايضا نظن أن الالات التى اخترعناها بعقولنا و صممناها بأيدينا مصممة لأن الداروينية توهمنا بذلك؟ لقد تعجب الفيزيائى الحاصل على نوبل يوجين وجنر من قدرة الرياضيات على العمل كلغة لكتابة و توصيف الفيزياء و على نفس المنوال فان قدرة الهندسة على توصيف الأحياء و الات الخلية مثيرة للدهشة و التساؤل [و كما يقول عالم الأحياء الجزيئية مايكل دنتون ”كل سمة من سمات تقنياتنا المتقدمة ممثلة بنظائرها فى الخلية...ان كثير من مصطلحاتنا التى نستخدمها لوصف ذلك الواقع المذهل فى الخلية مستمدة من عالم التقنية فى الجزء الثانى من القرن العشرين“و لعل من الأمثلة العجيبة على هذا الأمر ما يسمى Bacteriophage T4 و هو من الفيروسات المهاجمة للبكتيريا و الذى تم مقارنته بمركبات الهبوط على القمر lunar lander اذ له ثلاثة أرجل ليهبط بها على الخلية كما يمتلك مثقب لثقب الجدار و الية حقن و مولد طاقة حتى يعمل كل هذا و طبعا كل جزء من هذه الأجزاء الضرورية لهذه المركبة البيولوجية متكون من عدة أجزاء ضرورية تماما كمركبات فضاء ناسا]
المبحث الثانى : صعود جبل الاحتمالات البعيد [كشف تلبيس رتشارد دوكنز]
يحاجج الدراونة أعداء التصميم مثل رتشارد دوكنز بأن تصميم الأنظمة الأحيائية المتوهم فى زعمهم ليس مستحيلا من قبل الصدفة لأن التطور الداروينى يصعد جبل الاحتمالات خطوة خطوة فيقسم الاستحالة الى خطوات ضئيلة جدا تدخل كل منها فى حيز الاحتمال و سنبين فى هذا الفصل استحالة هذه الفرضية [من منظور معلوماتى و احتمالى بحيث يضاف الى ما وضحناه سابقا من عدم وجود خرافة الخطوات الصغيرة الداروينية فى السجل الأحفورى بل وجود الظهور الانفجارى المفاجئ لأنماط الكائنات الرئيسية. أيضا قبل أن نبدأ ننوه الى أن وجود سلم تدريجى من الطفرات المفيدة يربط البساطة بالتعقيد و يتشعب عبر كل الكائنات الموجودة فرضية مضمنة فى نظرية التطور اثباتها الوحيد حتى الان هو الأسهم التى يرسمونها بين الكائنات و مع ذلك يقفز التطوريون عادة فوق اثبات وجود هذا السلم التدرجى ابتداءا] يحاول الدراونة الترويج لأن الزمن هو البطل الحقيقى للتطور فباعطاء الأمر زمن كافى يغدو المستحيل ممكنا و بما أن الكون عمره بلايين السنين و يحتوى على عدد هائل من الكواكب يمكن أن يحدث فيه أى أمر بالصدفة نظرا لضخامة الموارد الاحتمالية. يأتى هنا المثل الداروينى الشهير الخاص بالقردة التى ستكتب أعمال شكسبير بالصدفة بالضرب على أزرار الالة الكاتبة.
قام الفيزيائى سيث لويد Seth Lioyd واضع نظرية الحوسبة الكمية Quantum Computing بمعهد ماساشوتس للتقنية MIT بحساب القدرة الحوسبية للكون Computational Capacity of the universe بناءا على كامل طاقته و عمره لتصل حساباته الى 400 بت من المعلومات كحد أقصى أى جملة مكونة من 82 حرف
See Seth Lloyd, “Computational Capacity of the Universe,” Physical Review Letters 88(23) (2002): 7901-7904. For a popularization of this work, see Seth Lloyd, Programming the Universe: A Quantum
Computer Scientist Takes on the Cosmos (New York: Knopf, 2006)
[قارن هذا بشفرة الحمض النووى الرهيبة المكونة من ملايين الحروف من المعلومات أو حتى بقصيدة واحدة لشكسبير] بل انه فى سنة 2003 تم عمل برنامج لمحاكاة فرضية قردة شكسبير Monkey Shakespeare simulator و قد وصل هذا البرنامج الى انتاج 24 حرف من أحد أعمال شكسبير فى محاكاة زمنية قدرها 2738 تريليون تريليون تريليون سنة/قرد [و هذا أكبر من عمر الكون بكثير و يختلف قطعا عن النصب الذى قام به رتشارد دوكنز عندما استعان ببرنامج حدد له جملة صغيرة سلفا كهدف مسبق و التطور لا هدف مسبق له يسعى الى تحقيقه بل يراكم التغيرات الصغيرة النافعة و لا يوجد شئ اسمه حرف نافع خارج سياق وجود جملة تم تصورها مسبقا فى ذهن مصمم البرنامج حتى يقيس عليها الحرف الصحيح/المفيد من الخاطئ و جعل البرنامج يحذف النتائج الخاطئة التى لا توصله الى الهدف و لا يكررها حتى يضيق النطاق الاحتمالى بينما حذف الانتخاب الطبيعى لطفرة لا يعنى عدم تكرارها مجددا ناهيك عن أنه أصلا يختار الفائدة الانية و لا يرى تصميم مستقبلى يعمل من أجله. كل هذا الى جانب أن تثبيت النتيجة الصحيحة لا يتم فى الطبيعة الا عبر تكاثر الكائن حتى تحل ذريته محل النوع القديم و هذا وحده يمكن أن يقاس بملايين السنين لبعض الكائنات. ان ما فعله دوكنز حرفيا هو ما يحاول اثبات عكسه و هو أنه وضع هدفا للتطور يحاول الوصول اليه. لقد علق عالم الرياضيات ديفيد برلنسكى ساخرا على هذا الأمر بعد أن فند عدم تشابه ما فعله دوكنز مع الاليات التطورية بل تشابهه مع التصميم الهادف قائلا أن ظهور اليات التصميم الهادف فى محاولة دوكنز هو ما يسميه عالم النفس فرويد بخروج المكبوت
David Berlinski. "The Deniable Darwin". Commentary. June 1996. p. 19-29
و طبعا كل هذا دون أن نتتطرق الى أن الطفرات و الانتخاب الطبيعى كثيرا ما يقودان الى الحذف و التدهور و الاتلاف على المدى البعيد لتحقيق منفعة انية و هو ما سنناقشه لاحقا ان شاء الله فى أيقونات التطور]
[من الأمثلة الأخرى على ذلك أيضا قيام الفيلسوف الملحد الذى امن اخر حياته أنتونى فلو بحساب احتمالى فى لحالة تحويل كل ذرات الكون لمعالجات حاسوبية processors يقوم كل منها بمليون محاولة فى الثانية منذ لحظة الانفجار الكبير الى يومنا هذا أى لمدة تقترب من 14 مليار سنة فان عدد المحاولات التى ستقوم بها 10 أس 90 فقط مقارنة مقارنة ب10 أس 690 محاولة مطلوبة لكتابة قصيدة شكسبيرية واحدة من 488 حرف بشكل عشوائى فما بالك بشفرة من 200 ألف حرف مطلوبة لبدء الحياة؟ قارن كل هذا الان بمعادلات المستحيل الرياضى التى حسبها العلماء بضرب عدد جسيمات الكون 10 أس 80 فى عدد التغيرات الممكنة فيزيائيا للجسيم فى الثانية 10 أس 45 فى عدد الثوانى منذ نشأة الكون 10 أس 25 ليكون أى أمر يتطلب موردا حتماليا أكبر من 10 أس 150 هو مستحيل الحدوث فى كوننا لأنه ببساطة لايمتلك جسيمات و ثوانى و تغيرات كافية لاحداثه...ان هذه البراهين الرياضية الفيزيائية الجازمة هى رد قاطع على محاولات الدراونة للتهرب فلسفيا من الزامات نظرياتهم بالاختباء خلف عاملي الزمن و الصدفة]
[و بعيدا عن كل هذه الحسابات المعقدة يرجى ملاحظة التالى...لا أحد عندما يقرأ كتابا أو يتعامل مع برنامجا حاسوبيا يفترض أنه فى مكان ما و زمان ما لابد أنه كانت هناك الاف أو ملايين الصدف أو أخطاء النسخ التى أنتجت هذا الكتاب أو هذا البرنامج...فقط يتم استدعاء هذا التخريف عند الحديث عن الحمض النووى و أدلة التصميم فى الكون ثم اعادته الى مزبلة الأفكار التى أتى منها و العودة الى العلم و العقل اللذان يقولان أن التصميم له مصمم و أن التشفير له مشفر و أن المعلومة مصدرها الذكاء بمجرد الانتقال الى أى حديث اخر حتى لو كان عن ما هو أقل تعقيدا!!!]
برغم ذلك يستمر الدراونة فى ترويج الوهم فيكتب دوكنز أن الأحياء هى دراسة الأشياء المعقدة التى تبدو لأول وهلة أنها مصممة و يكتب فرانسيس كريك "على علماء الأحياء أن يتذكروا دوما أن ما يرونه ليس مصمما و أنه نشأ بالتطور" [نعم مستر فرانسيس عليهم أن يذكروا أنفسهم باستمرار لأن كل شئ يصرخ فى وجوههم بالعكس - وذكر ديتكر شاميرز أنه بينما كان يتأمل ف أذن ابنته ورأى تعقيدها ودقة تركيبها جال ف ذهنه بأن هذا التعقيد وتلك الدقة لا بد لها من صانع، ولكنه طرد هذا الوسواس حتى لا يضطر إلى الإيمان بالخالق كما يذكر عن نفسه!!، ويعلق الدكتور تامس ديودباركس على هذه الحادثة قائلا: "إنني أعرف عددا من أساتذتي فى الجامعة، ومن رفقائي العلماء الذين تعرضوا مرارا لمثل هذه المشاعر، وهم يقومون بعمليات كيمياوية وطبيعية ف المعامل] كل هذا لتكذيب بديهيات الحدس و العقل و الاحصاء فهل فعلا يمكن أن تنجح الداروينية فى تخطى العقبات و انشاء التصميم حتى نكذب الحدس و العقل و الرياضيات؟ فلنرى
المبحث الثالث: عقبات التطور
من أجل تنسيق التغيرات التطورية اللازمة للتعقيد غير القابل للاختزال تواجه الالية الداروينية بلاغائيتها و تركيزها على انتخاب المنفعة الانية المتعلقة بالبقاء و زيادة النسل عددا من العقبات (فهى مثل المقاول الذى لا يرى الا اللحظة الحالية فبنى الجدار لمنفعته الانية لمنع اللصوص من دخول الأرض قبل الأساس الذى لا منفعة انية له فانهار البناء كله) و هذه العقبات يمكنتلخيصها فى النقاط الاتية:
1-التوفر: وجود الأجزاء المكونة للنظام المعقد حيث يجب تكوين البروتينات الجديدة المعقدة بالتصميم الشكلى والطى الفراغى protein folding الملائم فى أنظمة أخرى بصدف متتالية تعدل البروتينات الموجودة دون افساد الأنظمة الأصلية بمعجزة ما أو بصدف اضافية تؤدى الى وجود نسخ مكررة من الأنظمة الأصلية لتعوض فسادها
2-التزامن: حيث يجب أن يتم توفير كل الأجزاء المطلوبة فى نفس الوقت و للتزامن مستويين: الأول تطورى فانتاج جزء قبل بقية الأجزاء سيتركه عرضة لطفرات أخرى تفسده ما لم نضيف المزيد من الصدف لتوقف تطفره بعد بلوغه الصورة المطلوبة و الثانى جنينى أثناء نمو الكائن من البيضة الملقحة الى طور النضج حيث يجب أن يأتى انتاج مواد البناء فى الوقت الصحيح [بالصدفة طبعا] و الا مات الجنين أو تشوه [ملحوظة: فى حال محاولة الداروينى التفلت من المستوى الجنينى بأكذوبة ارنست هيكل recapitulation أن النمو الجنينى يعكس التطوريرجى مراجعة الفصل الخامس لاثبات كذب و تزوير هيكل]
3-التموضع: يجب أن تتجمع البروتينات المطلوبة من أماكنها السابقة فى أنظمتها الأصلية فى موقع تصنيع الالة البروتينية الجديدة و هذا يتطلب مستويين من الصدف – الأول صدف تكرار نسخ الموروثات التى تحتوى على الالية الأصلية لصناعة بديل لها و صدف نقل البروتينات المطلوبة من المكان القديم الى الجديد.
4-التفاعلات المتداخلة: اذا كان موقع البناء مفتوح على الكثير من البروتينات لتنسيق النقطة السابقة فان أى بروتين اضافى خارج قائمة البروتينات المطلوبة يتم نقله اليه سيتداخل مع عملية البناء و يفسدها لكن الصدفة الداروينية طبعا دقيقة جدا فى أعداد و أنواع البروتينات التى سيتم نقلها الى موضع تكوين الالية الجديدة
5-توافق الأجزاء: ان البروتينات بتصميمها المعقد ثلاثى الأبعاد و طياتها و ثنياتها تصبح مثل قطع بازل puzzle ثلاثية الأبعاد و لكى يتم تعشيقها معا بالشكل الصحيح يجب أن تتوافق أسطحها و تصاميمها ليتم التركيب كما يجب أن تتكامل وظائفها حتى تنجح اعادة الدمج و التوظيف فى النظام الجديد. يمكننا تسمية هذا بالتصميم المشترك حيث يتم نصميم كل أجزاء النظام و البروتينات بشكل يسمح باستخدامها فى أشياء أخرى مستقبلا و هو ما لا تستطيعه اليات صدفية لاغائية عمياء فمثلا عندما يزعم الدراونة هو نتيجةاندماج نظام الحقن TTSS مع أهداب البكتيريا متجاهلين نشأة و تعقيد كلا منهما فبالاضافة الى ما سقناه فى الفصل السادس لتفنيد هذا الزعم يمكن أيضا دحضه تجريبيا بالنظر الى الطفرات المطلوبة ليتوافق النظامان معا
6-ترتيب التجميع: لا يكفى أن تتجمع الأجزاء فقط فى الموقع بل يجب أن يتم تجميعها و تركيبها بالشكل و الترتيب الصحيح للوصول الى التصميم النهائى [مع ملاحظةوجود بروتينات للتجميع و هو ما يزيد الأمر تعقيدا. بل اننا يمكننا خلط مكونات الخلية الجاهزة فى معمل و مع ذلك لا تدب فيها الحياة أو باللغة المادية لا تعمل.]
يمكننا صياغة العقبات السابقة احصائيا على هيئة معادلة رياضية لتقدير احتمال ظهور التعقيد بالاليات الداروينية فمثلا يمكن دراسة توافق أجزاء الأنظمة الموجودة و مقدار قابليتها للخلط و التشويش تجريبيا أو حساب احتمال حدوث الطفرات المزعومة لجعل هذه الأنظمة تتوافق [كتوافق TTSSمع أهداب البكتيريا لصناعة السوط البكتيرى] و المدد الزمنية اللازمة لتثبيت الطفرات فى تعداد سكان جماعة حية من نوع معين (تكاثر الكائن حامل الطفرة النافعة حتى تحل ذريته محل الجماعة الأصلية) ليصبح لدينا دليلا رياضيا على عجز الداروينية على انتاج تعقيد النظام محل البحث بدلا من الاكتفاء بسر سيناريوهات تطورية نظرية [و بأسلوب القصص المصورة comic books برسم صور بينها أسهم] لتفسير التعقيد كما يفعل الدراونة
Michael J. Behe and David W. Snoke, “Simulating Evolution by Gene Duplication of Protein Features that Require Multiple Amino Acid Residues,” Protein Science 13 (2004): 1-14
تعرف المعادلة المستخدمة فى هذه العملية الرياضية باسم Origination Inequality متراجحة الانشاء و من اللطيف أن الدراونة يعترفون بها بل و يستخدمونها فى الحالات البسيطة عالية احتمال الحدوث للتدليل على الداروينية [و فى هذا أصلا مصادرة على المطلوب لأن الهدف دراسة التعقيد و ليس دراسة البساطة ثم الاستهبال – عفوا أقصد الاستدلال بها على التعقيد] و لكنهم يتجاهلونها عندما لا تؤيد نتائجها نظرياتهم و يتهربون بأنه بالتأكيد هناك سيناريوهاتتطورية تفصيلية لا نعرفها لذلك لم نتمكن من حسابها ملقين بذلك عبء ايجاد تفاصيل هذه السيناريوهات التطورية الممكنة كلها على المخالف ليدحضها دون أن يكلفوا أنفسهم عناء الاتيان بهذه السيناريوهات و تفاصيلها للتدليل على مزاعمهم. هذه الازدواجية فى معايير علم الأحياء التطورية فلسفة و ليست علما و تمنح الداروينية حصانة غير عادلة و هى ازدواجية تعسفية تعود الى داروين نفسه الذى قال أن نظريته يظهر خطأها بظهور عضو معقد يستحيل انتاجه بتعديلات كثيرة طفيفة متتالية دون أن يثبت أصلا أن هناك عضو ظهر بهذه الطريقة بل ألقى العبء على كاهل المعارض أن يأتى بكل السيناريوهات الممكنة و ينفيها!!!
[ملحوظة: فى هذا الفصل يقر الكتاب للدراونة بحجة مقاومة البكتيريا للمضادات الحيوية على أنها نموذج عالى الاحتمال منخفض التعقيد للتطور الصغروى بينما فى كتاب تفنيد أركان الداروينية الصادر لاحقا يفند دكتور جيرى برجمان حجج الدراونة المبنية على مقاومة البكتيريا للمضادات الحيوية مثبتا أنها اما انتخاب طبيعى دون تطور و اما تدهور و انتكاس...للمزيد من التفاصيل يرجى مراجعة تفنيد أركان الداروينية ]
المبحث الرابع: مستوى التعقيد
يحاول الدراونة عادة الاختباء خلف التعقيد الشديد صعب القياس للتفوه بأى هراء ثم ادعاء أنهم فسروه و بما أن تفسيرهم غير قابل للقياس فهو غير قابل للدحض فمثلا من الخرافات الداروينية الشائعة أن التطور قادر على تفسير عيون الفقاريات لكن حقيقة ما فعله الدراونة هو الاتيان بالأعين مختلفة مستويات التعقيد من الطبيعة ثم ترتيبها من الأقل تعقيدا الى الأكثر ثم رسم أسهم بينها و سموا هذا تفسيرا. لم يقدموا أى سيناريو واقعى أو نموذج تطورى تفصيلى لهذه القصص المصورة الطريفة التى يرسمونها من حيث تغير الشفرة الوراثية و النمو الجنينى و التوصيل العصبى لكن كعادة الدراونة ألقوا بعبء النفى على المخالف دون الاتيان بالاثبات أصلا [لقد رسمنا مجموعة من الصوربجوار بعضها...بل ووضعنا بينها أسهما كذلك ماذا تريدون منا أكثر منذلك...كم أنت رائع أيها العلم!!!]
بمثل هذه الطريقة عندما يكون تعقيد النظام جبارا لدرجة تجعل حساب احتمالات تطوره صعبا جدا يتهرب الدراونة من الزامات هذا التعقيد الداحضة لمزاعمهم بزعم عدم قابليتها للحساب لذلك كان مشروع مايكل بيهى هو تحديد أنظمة معقدة بدرجة معقولة تجعل حساب احتمالات تطورها ممكنا حتى يغلق على الدراونة باب التهرب.
يمكننا البدء بتقدير احتمال تطور بروتين وظيفى واحد الى اخر من خلال التغيرات الطفيفة المتتالية التى تتبناها الاليات الداروينية و التى ستغير شفرة الحمض النووى للبروتين خطوة خطوة صانعة بروتينا معدلا فى كل خطوة. يجب أن تكون كل سلسلة أحماض أمينية وسيطة قابلة للطى الفراغى و التشكل ثلاثى الأبعاد الى تصميم يقوم بوظيفة حتى يستبقيها الانتخاب الطبيعى. ان طى البروتينات من سلسلة أحماض أمينية الى شكل ثلاثى الأبعاد أمر مفصلى جدا فى تحديد وظيفتها و قابليتها للتفاعل مع بروتينات أخرى لذلك فهو غير قابل للتطور التدريجى الداروينى فاما أن يكون الطى و التصميم الشكلى صحيحا أو لا يكون لا يوجد أمر وسط أو مراحل تدريجية.
[و لكى نقدر أهمية الأمر يجب أن نفهم أن سلسلة الأحماض الأمينية التى يتشكل منها الانزيم لا فائدة منها حتى تتشكل فى شكل محدد. هذا الشكل يجلب أحماض أمينية من شتى أرجاء السلسلة ليصنع منها ما يسمى الموقع النشط active site و الذى يثبت فيه الانزيم الجزيئات التى سيعمل عليها و يخلق بيئة خاصة لاتمام التفاعل الكيميائى المطلوب. و الى جانب هذا يجب أيضا الاتيان بأحماض أمينية أخرى من السلسلة تعمل كأذرع ميكانيكية لنزع أو تركيب بعض الأجزاء لاتمام المنتج النهائى أو لترتبط مع بروتين اخر صانعة بنية أكبر حجما و أكثر تعقيدا تتكون من مجموعة بروتينات متناسقة. ان موقع الارتباط binding site يجب أن يكون مناسبا هندسيا و كيميائيا للهدف - هندسيا أى من ناحية الشكل كما يناسب القفل المفتاح و كيميائيا من ناحية المكونات فيجب أن تكون الشحنات الموجودة فيه مضادة لشحنات الهدف لاتمام التجاذب و الترابط و الا لحدث تنافر و بعد اتمام الارتباط يأتى دور المحفز catalytic site للقيام بالعمل على الهدف. نعم نحن نتكلم عن الة صناعية بالمعنى الحرفة للكلمة تعلم ما تفعله و كيف تفعله و للتوضيح يكم رؤية مثال على عمل الانزيم هنا
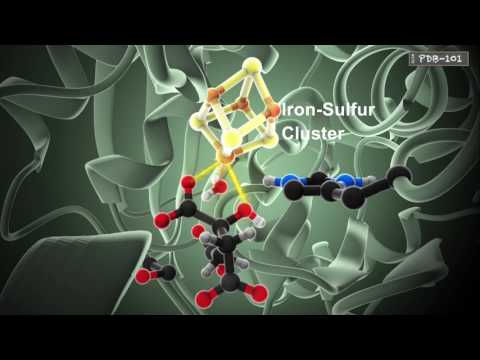
مثال اخر على التعقيد المحدد "روبوتات النقل فى الخلية"
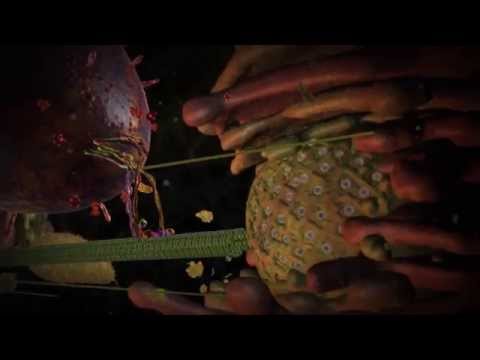
مثال من أبسط البروتينات بروتين الميوجلوبين مسؤول عن تخزين الأكسجين فى العضلات. يتكون الميوجلوبين (مثله مثل سائر البروتينات) من سلسلة أحماض امينية ذات صفات مختلفة فبعضها يتفاعل مع الماء و البعض الاخر ينفر من الماء و البعض ذو شحنة سلبية و البعض ذو شحنة ايجابية و غيرها من الخواص. تتفاعل هذه الخواص مع بعضها البعض و مع الماء فينجذب البعض فى اتجاهات بعينها و يندفع البعض فى اتجاهات أخرى و يأخذ البروتين شكل ثلاثى الأبعاد ملائم لاحتواء الأكسجين و لكنه يحتاج الى ما يشبه القفاز لينسك به الأكسجين هذا القفاز جزئ يسمى الهيم. بالصدفة طبعا يتواءم الهيم مع الجيب الذى يشكله تطوى البروتين فيدخل فيه كما تدخل اليد فى القفاز و يرتبط له و يثبت فى مكانه لكى يمسك به الأكسجين عند توافره و الهيم بدوره ملائم للارتباط بالميوجلوبين من جهة و للامساك بالأكسجين من جهة أخرى بسبب الصدف السعيدة طبعا. الهيموجلوبين بروتين مسؤول عن نقل الأكسجين فى الدم لذا هو الاخر يحتاج الى الارتباط بالأكسجين و بالتالى بينهما تشابهات كبيرة. طبعا بالنسبة لجهابذة التطور هذه التشابهات ليست أبدا بسبب القيود الوظيفية المتشابهة و العياذ بداروين و لكنها بسبب السلف المشترك. يتكون جزئ الهيموجلوبين من 4 سلاسل تلتحم ببعضها البعض بحيث يربط الجزئ 4 ذرات أكسجين فى المرة الواحدة.
اذن فيجب أن تكون لدينا الأحماض الأمينية الصحيحة فى التسلسل ثم يجب أن يأتى طى السلسلة protein folding ليضع الموقع النشط و الأذرع العاملة فى أماكنها الصحيحة و ان فكرنا بتحويل هذا بطريقة الخطوة خطوة الداروينية فلن ينتج لنا أى شئ لسببين. الأول أن نسبة التسلسلات الوظيفية و القابلة للتطوى ضئيلة جدا من بين كل التسلسلات الممكنة فاحتمال أن ينتج لنا التغيير العشوائى فى الحروف/الأحماض الأمينية شيئا فاعلا احتمال ضئيل جدا. ثانيا أن تحويل شكل معقد ثلاثى الأبعاد الى اخر يحتاج أن تكون هذه الطفرات متزامنة لا متوالية مما يضع عبئا أكبر على عامل الصدفة فتبديل حمض نووى واحد فى سلسلة كبيرة معقدة لن يحولها الى بروتين اخر. لاحظ أيضا أن بعض السلاسل البروتينية المعقدة تحتاج الى مساعدة لتحقق الانطواء بالشكل الصحيح فتأتى بروتينات أخرى تسمى شبرونات chaperones لتقوم بطيها و طبعا هذه يجب أن "تتطور" بالتزامن مع ما ستقوم بطيه و الا أصبح كلاهما بلا قيمة أى أننا أمام مجموعة متداخلة من خطوط الانتاج المعقدة يلقم كل منها الاخر]
حاول الدراونة الالتفاف على هذا الأمر بدراسة سلاسل أحماض أمينية قصيرة بسيطة أو بروتينات لا يتطلب عملها أن تأخذ شكل الالات الجزيئية المعقدة كبروتينات الارتباط ببلورات الثلج التى تمنع تجمد الدماء أو بروتينات ذات وظائف بسيطة أخرى لا تتطلب أكثر من الالتصاق بالهدف أو بعض التفاعلات الكيميائية البسيطة و فى هذا هروب من المطلوب ألا وهو دراسة التعقيد لا دراسة البساطة ثم تعميم نتائجها على التعقيد حيث تزداد صعوبة و تعقيد طى البروتينات و تشكيلها كلما زاد طول سلاسل الأحماض الأمينية و كلما زادت متطلبات و شروط الوظيفة لدرجة أن البروتينات الكبيرة المعقدة تحتاج الى بروتينات مساعدة لطيها طبقة طبقة فى تسلسلات محددة [و طبعا اليات المساعدة أيضا صدفة و لا تدل على القصد و لا التصميم بتاتا بل هى الداروينية] و يجب أن تتم خطوات الطى بترتيب محدد للوصول الى الشكل النهائى السليم المؤدى للوظيفة.
للرد على الدراسات المخادعة التى قام بها الدراونة على السلاسل القصيرة قام عالم الأحياء الجزيئية molecular biology دوجلاس اكس بدراسة على بروتين Beta-lactamase الذى يحمى الجراثيم من البنسلين و درس ميدان بروتينى من 150 حمض أمينى مثبتا عدم قدرة الاليات الداروينية على انتاجه و قد استخدم فى تجاربه تقنيات الحوسبة التطورية evolutionary computing التى تعتمد على معادلات و خوارزميات algorithms تطورية لنمذجة الداروينية حاسوبيا و خلص الى نتيجتين أساسيتين.
الأولى: احتمال تطور التطوى البروتينى المؤدى للوظيفة حتى لو كان-تنزلا-غير ثابت أو معاق هو 1 من 10 أس 64 و هذا-نكرر-وفقا لبرمجيات تطورية داروينية.
Douglas D. Axe, “Estimating the Prevalence of Protein Sequences Adopting Functional Enzyme Folds,”
Journal of Molecular Biology 341 (2004): 1298. Axe, “Estimating the Prevalence,” 1295.
الثانى: و هو الأهم لأنه لا ينطبق على هذا البروتين فقط و لكن على كل البروتينات و هو أن الطفرات و الانتخاب الطبيعى لا يعملان كفريق متناغم على المستوى الجزيئى البروتينى كما يتطلب التطور لأن التغير الطفيف فى البروتين لن يغير التطوى الخاص به و سيبقى البروتين المعدل بنفس التطوى و نفس الوظيفة و بالتالى لا يملك الانتخاب الطبيعى أى سبب لاستبقاء هذه الطفرة على حساب البروتينات التى لم تتطفر حتى تبنى عليها الداروينية طفرات و تعديلات طفيفة أخرى و هو ما يدل بوضوح على استحالة تطور أى بروتين معقد بيتا لاكتاماز أو غيره بالاليات الداروينية من تغيرات طفيفة و انتخاب طبيعى
[حاول الدراونة المراوغة مرة أخرى و نقد عمل دوجلاس أكس بزعم أن هناك بروتينات غير متطوية بشكل ثابت و هذه ربما تكون "حلقة انتقالية" الا أنهم تجاهلوا أن ما يسمونه حلقة انتقالية محتملة هو بروتين وظيفى و تطفره أو الاختلال فى مستوياته يقود الى حالات مرضية
https://evolutionnews.org/2018/04/protein-folding-and-the-four-horsemen-of-the-axocalypse/
https://evolutionnews.org/2018/02/losing-the-forest-by-fixating-on-the-trees-a-response-to-venemas-critique-of-undeniable/
Peter E. Wright and H. Jane Dyson "Intrinsically Disordered Proteins in Cellular Signaling and Regulation" Nat Rev Mol Cell Biol. 2015 January ; 16(1): 18–29
و الأهم من ذلك وفقا لنفس الورقة فان ما يزعمون أنه بروتين غير متطوى هو فى الواقع بروتين يحتوى على عناصر تمييز small recognition elements that fold upon partner binding تجعله يتطوى عن التقاط الهدف المناسب أى أنه يخضع لتطوى مشروط Conditional Folding فهو يمتلك تطوى البروتين العادى بالاضافة الى الية اضافية لالتقاط الشروط المطلوبة للتطوى و تفعيله أى أنه دليل على تصميم أعقد و تعقيد أكثر تحديدا من البروتين العادى...و فى دراسة أخرى وجد فريق الباحثين أن البروتين يحتاج الى تموضع أحماض أمينية بشحنات متنافرة فى مواضع بعينها لمنع البروتين من التكتل ختى تحين اللحظة المناسبة و يتم تعديل الشحنات للسماح بالتجاذب بين سلاسل البروتين (انظر الشكل رقم 5 فى الورقة)
Our results collectively show that a subtle balance between electrostatic repulsion and interchain attractive interactions regulates CAPRIN1 phase separation and provides insight into how nucleotides, such as ATP, can induce formation of and subsequently dissolve protein condensates....At least 75% of IDPs contain both positively and negatively charged residues, with charge–charge interactions important in defining their physical and chemical properties and, in some cases, their propensities to phase separate...electrostatics play a central role in modulating the biological functions of this protein, and measurement of electrostatic potentials at each site along its backbone, as reported here, provides an opportunity to understand in more detail the important role of charge in this system.
Yuki Toyama et al., "Mapping the per-residue surface electrostatic potential of CAPRIN1 along its phase-separation trajectory" PNAS Vol. 119 | No. 36 , August 30, 2022
أى أن هذا النوع من البروتينات هو الاخر يحتاج الى تسلسل دقيق و تموضع أحماض أمينية بعينها فى مواقع بعينها للحفاظ على حالة التطوى الشرطى اللازم للقيام بوظيفته...من جديد تثبت الأبحاث أن هذه ليست حلقات انتقالية نحو أى شيء و أنها تحتاج الى تسلسل دقيق هى الأخرى ]
منقول من ملخص كتاب تصميم الحياة - د. ويليام ديمبسكي ود. جوناثان ويلز



تعليق